To locate and comment on Tallulah Pellissier-Lloyd's lichens-on-lampposts post, go to her blog.
Tuesday, October 19, 2021
Growing on Everything but Little Understood: The Strange Ecology of Lichen
Monday, October 18, 2021
The Least Dangerous Game: Tracking the Habitat Use of Squirrels on Campus
Fox squirrels (Sciurus niger) are a common sight here in Grinnell and much of the midwest. Indeed, fox squirrels seem to have made themselves quite comfortable in what is, ostensibly, an urban landscape here on campus. McCleery et al. 2007 takes a look at a similar situation on Texas A&M University Main Campus, tracking squirrels to examine their use of habitat in an urban environment. As more rural landscapes are urbanized, it is imperative to learn how squirrels, among other city-dwelling animals, interact with the urban environment so these areas may be managed to encourage, or discourage, their presence.
To track the habitat use of fox squirrels, the campus was delineated into four main habitat substrates: pavement, building, grass, and tree canopy. Prior to the study campus squirrels were trapped and fitted with a collar and radio transmitter. These radio-collared squirrels were tracked from September 2003 to August 2005, with the squirrels being located 2-3 times a week at random intervals. When located, their habitat substrate and activity were recorded into the GIS database. With these methods, the researchers were able to analyze habitat use according to both spatial and temporal scales. These scales included point-study area, range-study area, and point-range. Tree selection and squirrel activity were also examined.
Figure 2 (below) shows point-study area selection ratios of urban substrates (calculated by taking observed habitat use divided by expected habitat use for each substrate and season). Similar figures were used to depict the results from the other spatial scales. They found that fox squirrels strongly select for tree canopy regardless of scale and activity level. Conversely, fox squirrels generally avoid pavement but do not exclude it from the core areas of their ranges. It was also observed that fox squirrels used buildings both as a substrate and daytime refuge during the winter and spring. Tree selection analysis indicates that the squirrels generally avoided pines, bald cypress, and arborvitae, but selected for live oaks. During periods of activity squirrels selected for trees with greater canopy area and diameter at breast height.

These findings suggest that squirrels select for substrates similar to those they use in the wild, while adapting to, exploiting, or simply tolerating, features of the urban environment. This adaptability is likely what makes squirrels so successful in many urban areas, such as Grinnell College Campus. Our study takes a broad view of squirrel habitation on campus by comparing present-day nest distribution to that observed by a previous study. Our goal is to see how squirrel nest distribution has changed, if at all, since this study was conducted and investigate potential causes. Specifically, we want to examine if derecho damage sustained by trees on campus has affected squirrel nest distribution.
References
McCleery, R. A., Lopez, R. R., Silvy, N. J., & Kahlick, S. N. (2007).
Habitat use of fox squirrels in an urban environment. The Journal of wildlife management, 71(4),1149-1157.
https://grinnell.idm.oclc.org/login?url=https://www.proquest.com/scholarly-journals/
habitat-use-fox-squirrels-urban-environment/docview/234183395/se-2?accountid=7379
Saturday, October 16, 2021
"Leave Nothing but Footprints": Undergrowth Vegetation and Visitor Pressure
In the process of researching understory vegetation for our urban ecology project, I came across a study by Sikorski et al. that was interested in the effects of visitor pressure on the understory vegetation of forests. Prior to this study, other research has been done on the factors that contribute to an urban park’s species diversity of understory vegetation. The factors associated with the presence of visitors, also known as visitor pressure, are assumed to be the greatest contributor to declines in species diversity. However, no one had compared how species diversity decreases in trampled parks in ancient vs. recent forest stands. The authors of this study hypothesized that understory vegetation in park forest stands exposed to visitor pressure will decrease in species diversity.
The researchers analyzed the effects of visitor pressure on vegetation at thirty-seven parks in Warsaw, Poland. The data was split into three groups: recent forest stands with public access to undergrowth (PR), recent forest stands with restricted access to undergrowth (RR), and old forest stands with restricted access to undergrowth (OR). Older forest stands have been maintained for over 150 years ago and recent forest stands were planted approximately 60 years ago on previously non-forest environments. In each of these forests, the trunks of randomly selected trees were used as a central point to sample from 3 1x1m quadrates that measured vegetation composition, soil samples, and other habitat parameters.
Recent forest stands with public access to undergrowth (PR) were found to have fewer native species and fewer species overall in comparison to restricted access area species (RR and OR). However, the biggest finding was that RR stands had 30 times more species richness than PR stands. Additionally, PR stands exhibited features that hamper the growth of understory plants like higher pH, more soil compaction, a greater percentage of light reaching the ground, more bulk density, and a greater difference in moisture.
The urban ecology project I’m working on with my group also focuses on the understory vegetation of trees, with the biggest difference from Sikorski et al.’s work being that our focus is on small groups of trees rather than forests. We’re also limiting our focus to measuring the radius of vegetation, the distance between trees in a group, and the understory vegetation species. With the paper’s findings on the effects of visitor pressure in mind, I think close together communities of trees will be better protected from foot traffic and have greater species richness.
Reference:
Sikorski, P., Szumacher, I., Sikorska, D. et al. Effects of visitor pressure on understory vegetation in Warsaw forested parks (Poland). Environ Monit Assess 185, 5823–5836 (2013). https://doi.org/10.1007/s10661-012-2987-0
Friday, October 15, 2021
Ants on Trees: What Trees Do Ants Find Attractive in Urban Areas?
Urbanization
causes a lot of disturbance in the habitats organisms occupy and cause changes
in species composition in a certain location. Foreign species may be introduced
into a new locality which can form a potential habitat with mixed native and foreign
species. General fragmentation of habitat due to urbanization may also mean
that these potential habitats can act as refuge for many organisms, including
ants. This also offer some challenges to the foraging ants, as the resource
structure can be changed. The types of tree species can be especially
influential in ant distribution, as ants can be closely associated with their host
plant species or switch to another host if there is more honeydew availability
(MacArthur-Waltz et al., 2021).
MacArthur-Waltz
et al. (2021) looked into the preference of trees of ant species in California
on a college campus. In addition, they also observed if the health condition of
trees, seasonality and time of the day mattered to ants. They found ants are
overwhelmingly occurring on ever-green tree species in all seasons. Deciduous trees
were not as popular. This was the trend in both native and non-native tree
species, but ants significantly preferred native ever-green trees more. Ants
also preferred healthier trees to some extent, although trees with less rigor
ratings also had ants. The authors looked for evidence of resource partitioning
among different ant species. They found that different species are most active in
different seasons of the year and in different time of the day, suggesting that
could be a temporal resource partitioning between different ant species. However,
it is unclear if different ant species partitioned the same food resources.

This
article is closely related to our group’s project. It hints that we might be
able to find similar patterns in which ants preferred native tree species. It’s
doubtful if we can observe the same preference for evergreen trees. In the study
by MacArthur-Waltz et al. (2021), the ants were active all-year round and the
available evergreen tree species were quite different from the situation in Iowa.
However, the authors’ suggestion that ants’ preference for evergreen was due to
potentially more consistent resource availability might hold true for our study.
Ants preferring healthy trees might also be due to a similar reason. In our study
site, where some trees have been damaged sometimes severely by the derecho, this
could mean that ants would stay away from damaged trees.
Reference:
MacArthur-Waltz,
D. J., Nelson, R. A., Lee, G. and Gordon, D. W. (2021). Tree Preference and
Temporal Activity Patterns
for a Native Ant Community in an Urbanized California Woodland. Journal of Insect Behavior, 34, 211-222.
Enlightening Lichens
Air pollution has become a topic that is increasingly researched and talked about, and understandably so. It can have catastrophic effects on ecosystems, especially urban ecosystems, therefore negatively impacting human health as well as the surrounding environment. A symbiotic relationship between fungi and algae, lichens are key organisms when it comes to indicating air quality, pollution levels, and overall ecosystem health (National Park Service). Because they collect all of their necessary nutrients from the air, they are particularly sensitive to pollutants. The absence of lichens can indicate poor air quality or the presence of particularly toxic pollutants. Not only are they wonderful for indicating the health of an ecosystem, but they also serve as food to many organisms and clean the air, making them especially beneficial to the places they are found (National Park Service).
Despite lichens being ecologically important, not much is known about them and not much research has been done since the late 1900s/early 2000s. In 1997 a study was done in the Aniwaniwa River Valley of North Island, New Zealand, by Green et al. They looked at net photosynthesis (NP) of nine species of rain forest lichens in response to differing levels of light - from shaded areas in the forest through open ground sites, and NP of Pseudocyphellaria coronata in response to water content (WC) at constant light. They found that species found in the open areas had much higher light saturation (the amount of light needed for max net photosynthesis) than species found in and on the edge of the forest, and that as light increased, NP increased quickly until it reached light saturation when it tapered off and remained constant (figure 2). The authors noted that lichens dry out quickly once in the sun, so the lichens in the open habitat could photosynthesize at high light levels, but they were only active for 50% of the measurements. They suggest that the structure of the open habitat lichens (fruticose) may help by providing shade for itself. For P. coronata (found in half shade), they found that at low WC and high WC, NP and light saturation decreased with NP decreasing at many light amounts measured (figure 6). This suggests the importance WC plays for the metabolic and photosynthetic abilities of the lichen.
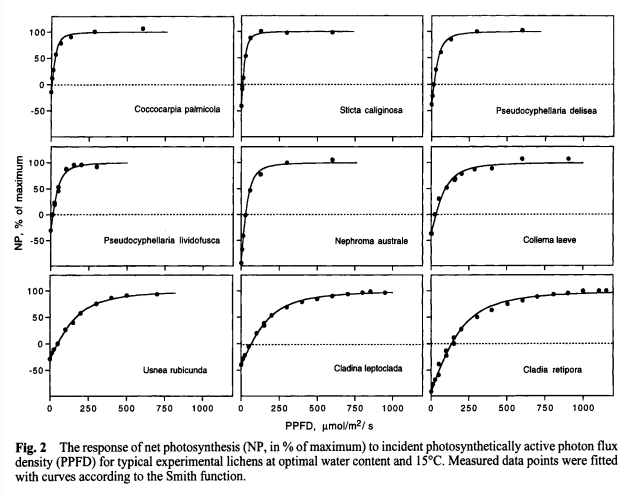
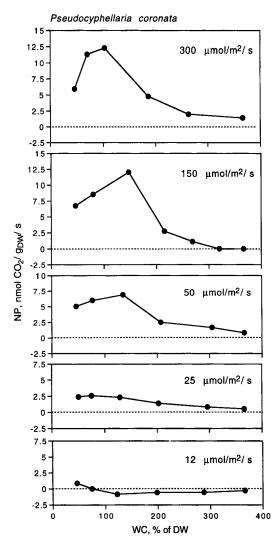
Figure 1: A (on the left) figures from Green et al. 1997 showing the response of NP to incident photosynthetically active photon flux (PPFD or amount of light as I have been referring to it) at optimal WC and 15C. B (on the right) figures from Green et al. 1997, showing NP in response to differing WC at constant light levels (also measured in PPFD).
Continuing on from this paper, Bjorn, Tallulah, and I are looking at lichens on the lamp posts of south campus. One aspect of the lichens we are measuring is the directionality distribution (north, east, south, west). Because of the dependency of lichens on light and water, as found by Green et al. (1997), the lichens we are looking at will want to be at light saturation, but also have an optimal WC. If our lichens are able to photosynthesize at a high light saturation and low WC, they may be found on the sunnier (south) sides of the poles, but if WC content plays as important a role it did for Pseudocyphellaria coronata, our lichens will likely be found on sides that spend less time in direct sunlight.
References:
National Park Service. (2018). Lichen - The little things that matter. https://www.nps.gov/articles/lichen-and-our-air.htm
T. G. A. Green , B. Büdel , A. Meyer , H. Zellner & O. L. Lange (1997) Temperate rainforest lichens in New Zealand: Light response of photosynthesis. New Zealand Journal of Botany, 35:4, 493-504, DOI: 10.1080/0028825X.1987.10410173
Many studies on wildlife populations focused on their
natural histories in pristine settings. However, our landscapes are
increasingly comprised of urban environments characterized by buildings, high
availability of food, water, and energy, and perhaps more importantly more
humans. Naturally, we would expect populations in urban environments to display
a different set of physiology, habitat selection, and behaviors than their
rural counterparts. Thus, it is important to study urban populations
systematically. McCleery & Parker (2011) looked at city squirrels.
Specifically, they investigated the influence of urban environments on fox
squirrel range overlap.

Photo from Grinnell Magazine by Grant Dissette '12
Squirrels are territorial. Studies have shown that squirrels
in the traditional rural settings are hierarchical and characterized by
extensive overlapping of ranges (Koprowski 1996). McCleery & Parker
examined the spatial overlap in squirrel ranges during different seasons. They
considered more overlap as hierarchical behaviors and less overlap to be
territorial behaviors.
The urban part of the study was conducted on the main campus
of Texas A&M University dominated by buildings, pavement, and manicured
exotic greases, while the rural part was on a private ranch of the University
of undisturbed hardwood forest and meadows with no little to no human activity.
Twenty squirrels (10 male, 10 female) from each site were trapped with
wire-cage traps and tagged with radio-telemetry collars. Their seasonal ranges
were recorded and calculated.
The results show a reduced seasonal range overlap by
squirrels on the urban site. The study also found that urban squirrels used
fewer nests in winter and in general fewer tree cavities and shelters. McCleery
and Parker believe that squirrels adjust their behavior to use spaces based on
their environmental conditions. They proposed that range boundaries might be
influenced by buildings and the increased habitat unevenness may help delineate
territories. They think cavities might be a limited resource on urban sites and
maintained for exclusive use, such as pregnant females.
In our campus ecology project, Sharene, Jocelyn, and I will
be examining a slightly different aspect of urban squirrel populations –
preferred nesting locations. We record the tree species where nests are found,
their coordinates, and the position of the nests in the tree. In addition, we
will also be comparing our data to a previous data set also collected by
Grinnell students and see if the preferences of squirrels have changed over
time.
Koprowski, J.L. (1996). Natal philopatry, communal nesting, and kinship in fox squirrels and gray squirrels. J. Mammal.77, 1006–1016.
McCleery, R. A., & Parker, I. D. (2011). Influence of the urban environment on Fox Squirrel Range Overlap. Journal of Zoology, 285(3), 239–246. https://doi.org/10.1111/j.1469-7998.2011.00835.x
Understanding Squirrel Distribution in an Urban Landscape: What are they nuts for?
Squirrels...they’re cute, funny, and sometimes confusing like when they hang out in the South Campus loggia or do other silly things (Image 1). Some might even say they’re Grinnell College’s unofficial mascot. Even though it’s hard to imagine campus without the squirrels, many of us might consider “natural” forests as their true home and find the study of these areas to be most relevant to understanding squirrels and their conservation. As Salsbury et al. (2004) notes, “Effective species management practices in an age of dwindling natural, undisturbed habitats, however, necessitate that populations embedded within urban and agricultural landscapes be examined” (pp.369-370). Salsbury et al. (2004) does just that by studying fox squirrels (Sciurus niger) in Indianapolis, Marion County, Indiana. To their knowledge, they were the first to research squirrel populations living within a “human populated landscape transected by numerous roadways and dominated by residential and nonagricultural commercial properties” (p.370).

In order to understand how fox squirrels are affected by urbanization and habitat fragmentation, Salsbury et al. (2004) studied six woodlots from 1.1 to 8.3 ha in size and conducted leaf nest and vegetation surveys. For the leaf nest survey, the researchers tried to find all of the leaf nests in each woodlot, and they documented tree species, tree diameter at breast height (dbh), tree height, nest height, nest position, and the presence of vines. When completing the vegetation survey in each woodlot, Salsbury et al. (2004) sampled trees from at least one 100 m transect in the center and at least one 100 m transect that was 5 m from the edge. They noted tree species, tree dbh, the distance between trees and the transects, and the presence of vines. They also estimated the density of shrub and herbaceous cover. Salsbury et al. (2004) found that the squirrels preferred certain tree species but that their preferences differed between woodlots. This suggests that squirrels do not necessarily favor oaks and that other factors beyond tree species are important to them. In addition, larger trees and higher locations in trees tended to house nests. At four out of six of the sites, the squirrels also appeared to prefer trees with vines. Furthermore, Salsbury et al. (2004) found more nests at the disturbed woodlots, which could potentially indicate that squirrels favor areas with dense shrub cover. Together, this research reinforces the idea that squirrels are versatile and that they might even be positively affected by urbanization.
In our campus ecology research project, Hang, Jocelyn, and I will be building upon this research by studying the distribution of squirrel nests around the Grinnell College campus (Image 2). We are recording the tree species that nests are found in and their coordinates as well as the amount and position of nests in trees. We will investigate whether the tree preferences of Grinnell squirrels are also flexible. In addition, we will compare our squirrel nest distribution data to previous student research to see whether the preferences of Grinnell squirrels have potentially changed over time. In particular, we are interested in whether nest distribution has changed more in the sections of campus that experienced greater damage by the 2020 derecho.

Reference
Salsbury, C. M., Dolan, R. W., & Pentzer, E. B. (2004). The Distribution of Fox Squirrel (Sciurus niger) Leaf Nests within Forest Fragments in Central Indiana. The American Midland Naturalist, 151(2), 369–377. http://www.jstor.org/stable/3566753
The Unclear Fate of Fungi: How Landscape Management Practices Influence Fungal Populations
With the increasingly rapid transformation of wild spaces to agricultural and residential land, the study of how to best conserve plant and animal populations has become a priority among ecologists. In the study of ecosystem conservation, one important group of organisms is often neglected: fungi (Heilman-Claussen). Despite their vital functions in the ecosystem as decomposers, nitrogen cyclers, and mutualists, little study has been performed on how fungi may be conserved, or even how endangered fungi species are—the status of most fungal populations is unknown, with red list assessments rare (Komonen et al). As forests are fragmented and cleared, the availability and variety of wood substrates are greatly reduced, potentially limiting the range of saprophytic species (Moose et al). The study of small-scale forest management effects can thus provide a useful window into the effects of environmental alteration on fungi at large.
In their study of an agricultural area in Central Finland, Juutilainen et. al examine the impacts of different land management strategies on fungi populations. They defined plots within land split into three categories—natural broadleaf herb-rich forest, forested pastures, and afforested fields (formerly non-forest areas transformed into forests by humans)—and searched every piece of deadwood within the plots for fungi. Out of 67,130 deadwood particles, they found 209 species of fungi, with the highest species richness occurring in natural herb-rich forests and the second highest occurring in afforested fields. Natural herb-rich forests had a significantly higher number of unique species than the other land types, likely due to the higher presence of woody debris with large diameters, such as snags and logs, which provide more substrate and create more microclimates. Forested pastures were less hospitable to specialist fungi due to lower deadwood presence and drier microclimates. Juutilainen et al.’s study reveals that deadwood diameter and microclimate availability are crucial in supporting fungal diversity and suggests that better deadwood management—such as leaving logs and snags in place—could benefit fungal populations in heavily managed areas.
In our study, like Juutilainen et al., we divide land into categories—unmanaged, heavily managed, and semi-managed—and take random quadrats from each category in order to examine how management practices and deadwood presence affect fungi populations at Grinnell College. Although the semi-urban forest communities of Grinnell are very different from the birch forests of Finland, they are similarly fragmented by human use. We expect deadwood and fungi presence to negatively correlate with the degree of management—in highly managed areas like the South Campus lawn, for example, large pieces of deadwood are removed, reducing available substrate for saprophytic fungi. Our study could shed light on how management practices can be used to support fungi at Grinnell.


Measuring forest plots in Wieman, a less managed woodland on campus.
Heilmann-Clausen, J., Barron, E. S., Boddy, L., Dahlberg, A., Griffith, G. W., Nordén, J., Ovaskainen, O., Perini, C., Senn-Irlet, B., & Halme, P. (2015). A fungal
perspective on conservation biology. Conservation Biology, 29(1), 61–68.
https://doi.org/10.1111/cobi.12388
Juutilainen, K., Mönkkönen, M., Kotiranta, H., & Halme, P. (2016). The role of novel forest ecosystems in the conservation of wood‐inhabiting fungi in boreal
broadleaved forests. Ecology and Evolution, 6(19), 6943–6954.
https://doi.org/10.1002/ece3.2384
Komonen, A., Sundström, L. M., Wall, A., & Halme, P. (2016). Afforested fields benefit
nutrient-demanding fungi: Afforested fields benefit fungi. Restoration Ecology,
24(1), 53–60.
https://doi.org/10.1111/rec.12282
Moose, R. A., Schigel, D., Kirby, L. J., & Shumskaya, M. (2019). Dead wood fungi in North America: an insight into research and conservation potential. Nature
Conservation, 32(32), 1–17.
https://doi.org/10.3897/natureconservation.32.30875
Mushrooms on the Defensive: Fungal Decline in Urban Versus Natural Areas
It is widely known that biodiversity is declining worldwide due to anthropogenic forces like climate change and habitat loss. The ramifications for ecosystem integrity are massive, and the challenge of stopping this downward slide is equally daunting. This decline is found across groups, from plants to birds to fungi, and is especially true in urban areas because of the transformation of previously available habitat. Often inconspicuous and sometimes seen as insignificant, fungi in fact play an important role in many ecosystems. They decompose dead organic matter and return nutrients like nitrogen to the environment that would otherwise stay locked in those dead organisms. There the nutrients can be used by plants and animals. They also form symbiotic relationships with plants and can behave as indicator species. Their population decline needs to be investigated to better understand how and why it is occurring and improve management practices.
Abrego et al. (2020) takes an innovative approach to quantifying this decline. They investigated fungal diversity in urban and natural areas, using DNA analysis of soil samples and aerial spore samples to assess the prevalence, number, specialization, and composition of different species. They sampled plots within larger areas representing urban and natural areas, in both the core of these areas and their edges, in or near five different Finnish cities. Measuring in multiple cities allowed the researchers to track how communities changed over larger distances than just those shown in the example areas below.
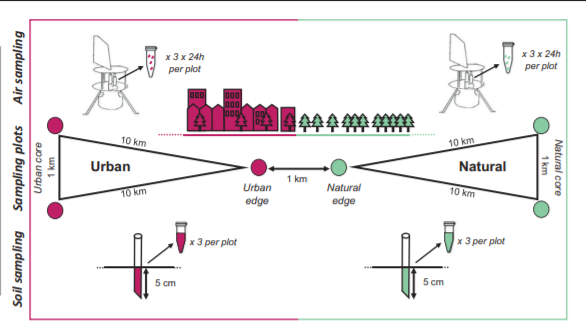
Figure 1. Image from Abrego et al. (2020) demonstrating the sampling points and the techniques used to sample from the air and soil. The tool at the top is a cyclone sampler, used for collecting aerial mushroom spores.
What’s interesting in what they found is that there wasn’t an overall difference in the total number of species between urban and natural areas. There were, however, more species per sample in the natural areas even though the overall number was the same. They also found many more natural specialist mushrooms than urban ones, and more in the air than the soil- both of generalists and specialists. Surprisingly, the air samples indicated a larger drop in diversity and abundance between natural and urban habitats than the soil samples. The researchers had thought that it would be the soil samples that would show lower abundance in urban habitats because of limited dispersal relative to wind-born spores. Instead of being homogenous over large areas, aerial fungal communities demonstrate a structure based on location.

Figure 2. Image from the article visualizing their results. S represents the predicted species richness per sample, while the colors represent the specializations of the fungi identified. Specialists were defined as species more than 10 times more prevalent in one habitat than another.
These distribution patterns are certainly influenced by management practices within these areas, a question we are currently studying in an urban ecology project on the Grinnell College Campus. We’re looking at how gradients of management across campus influence the presence of dead wood and the saprotrophic fungi that colonize it. Our study should provide insight into how our management practices influence habitat availability and population distribution for these important organisms.
These results symbolize the reduction in diversity that is happening across taxa and emphasize the need to conserve and protect natural areas. Fungi are important as decomposers and plant symbionts. They must be taken into consideration in any conservation planning to preserve the integrity of existing ecosystems and the future functioning of the planet.
Citation
Abrego, N., Crosier, B., Somervuo, P., Ivanova, N., Abrahamyan, A., Abdi, A., Hämäläinen, K., Junninen, K., Maunula, M., Purhonen, J., & Ovaskainen, O. (2020). Fungal communities decline with urbanization—more in air than in soil. The ISME Journal, 14(11), 2806–2815. https://doi.org/10.1038/s41396-020-0732-1
How do seeds travel when they're on an (urban) island?
As urban areas become increasingly prevalent, ecologists are turning to study the (previously unrecognized) natural aspects of these environments. Urban areas offer different resource limitations and selective pressures than rural or natural areas, so studying other communities in these environments can illuminate interesting ecological phenomena. One of the challenges that plant communities face in these conditions is seed dispersal. In a city-like environment, resources are more likely to be fragmented by concrete or asphalt, leading to isolated patches of growth. The question is, then, if this island effect changes the efficacy of a species seed dispersal mechanism, and if this is putting selective pressure on plant communities. Cheptou et al. (2008) investigated this phenomenon in their research on the evolutionary consequences of seed dispersal in the weedy, allogamous plant Cresis sancta.

Crepis sancta, http://www.freenatureimages.eu/plants/flora%20c/Crepis%20sancta/index.html
Since C. sancta produces both dispersing and non-dispersing seeds, the researchers were able to plant C. sancta in artificial patches throughout a city south of France, and measure how many dispersing and non-dispersing seeds were produced in a given season. Then, they were able to germinate the seeds from this first experiment in the field in the lab, and measure how urban plants compared in their seed dispersal profile to populations in “country-side-like” conditions. What the researchers found was that dispersing seeds had, on average, a 55% lower chance of landing in suitable habitat, which makes sense when considering the likelihood of a seed landing on concrete in a city. Non-dispersing seeds were, therefore, more likely to land on suitable ground. In a comparison of urban and rural populations’ progeny, the researchers also found that urban populations produced a significantly higher ratio of non-dispersing seeds. Genetic modeling showed that this shift in the ratio of seed dispersal was exceptionally quick, demonstrating that fragmented landscapes such as those found in cities put a lot of selective pressure on communities to produce more non-dispersing seeds.
Cheptou et al.’s research highlights the impact that fragmentation has on plant communities, even down to their ability to spread to other areas. Riana, Catherine and I are similarly interested in studying fragmentation of plant communities, but we are studying patches under spruce trees on Grinnell’s campus. The microhabitats under spruce trees may be impacted by the island effect, especially if the plants in question cannot disperse their seeds in the same way that they might in a non-fragmented area, as Cheptou et al.’s research suggests. We suspect that spruce trees in proximity to each other may have similar plant species profiles, while spatially segregated spruces will be fairly different. For example, a disturbance-tolerant plant like Plantago major that sends out wind-dispersed seeds might not be present in disturbed areas that we would expect it to be because of these added difficulties in seed dispersal. Granted, the conservation of P. major is not a concern among ecologists because it is weedy, but this research is relevant in understanding these patterns of dispersal in the plant community. Microhabitats under trees may prove to be a vital source of biodiversity in our increasingly urban human habitats.
References
Cheptou, P.O., Carrue, O., Rouifed, S. & Cantarel, A. (2008). Rapid evolution of seed dispersal in an urban environment in the weed Crepis sancta. PNAS, 105(10), 3796-3799.
Below the Branches: Fragmentation Impacting Understory Plant Communities
As humans continue to take control over an ever-increasing amount of natural and semi-natural spaces through acts such as deforestation and urbanization, we inevitably change the habitats of all the other organisms living there. Although organisms, including plants and animals, can persist in these severely altered landscapes, their populations are oftentimes quite fragmented. In light of this, Benítez-Malvido & Martínez-Ramos (2003) (found here) became curious about the ways that forest fragmentation in the Amazon Rainforest impacts the plant communities that live under these tree canopies, particularly as fragmentation alters community structure, reproduction, demography, and reduces biodiversity (Dubois & Cheptou, 2017).
To study the impacts of fragmentation, Benítez-Malvido & Martínez-Ramos (2003) isolated 11 different fragments of forest of either 1 ha, 10 ha, or 100 ha that were all located within cattle ranches, a major source of fragmentation and habitat destruction in the Amazon Rainforest. They also had access to a continuous forest that was still near agricultural lands, but not fragmented by it.
Benítez-Malvido & Martínez-Ramos (2003) found showed that species richness was significantly higher in the continuous forest compared to the fragments, and, in fact, the researchers were unable to record the entire richness of the continuous forest as it was so vast. Further, the overall trend shows that the smaller the plot, the lower the species richness. Surprisingly, the researchers also note that fragments that were closer together did not have greater similarity to each other than fragments that were farther apart. Instead, fragment size made the greatest difference for species composition. The only species that this did not hold true for was a tree species in the Burseraceae family that became dominant in all sites, regardless of size. The researchers suggest that traits such as high reproduction independent of the environment, successful germination, using generalist pollinators, stress-tolerant seedlings, and resistance to natural enemies may be the best indicators of why a species may still be able to flourish in fragmentation.

Although Iowa is not the Amazon (sorry to break the news..), these questions are still pertinent to our urban fragmented tree clusters and solo trees that harbor understory plant communities. Here on campus, our fragments are separated by regularly mowed grass and, further beyond, surrounded by oceans of agricultural fields. We have sampled under 41 Spruce trees thus far, and we are paying attention to the total area of the plant community under the tree, the plant community’s distance to the nearest spruce’s plant community, and keeping records of the species we see under each tree. Although we are not comparing our tree fragments to a continuous forest, we are still looking at similar trends as Benítez-Malvido & Martínez-Ramos (2003), including similarity of plant community composition based on distance from other communities and species richness based on location on campus and total area of the plant growth under the trees. Additionally, we are looking for general trends that give insight into which plants perform best under disturbance conditions, which we can take into account as some of the spruce trees have foot traffic and potential mowing under them, while others do not due to low branches creating protection. I am curious if we will discover similar results to Benítez-Malvido & Martínez-Ramos (2003), perhaps showing an importance of area under the trees for diversity and no significant difference based on proximity to other plots, or perhaps our “urban” Iowa understory plants may reveal different trends.

Riana Mulligan and Emma Thran befriending Spruce #22.
References:
Benítez-Malvido, J., Martínez-Ramos, M. (2003). Impact of forest fragmentation on understory plant species richness in Amazonia. Conservation Biology, 17(2), 389-400.
Dubois, J., Cheptou, P.O. (2017). Effects of fragmentation on plant adaptation to urban environments. Philosophical Transactions of the Royal Society: Biological Sciences, 372, 1-8.
Subscribe to:
Posts (Atom)